|
|
|
|
|
|
|
|
|
|
|
Само
совершенство
Наш рассказ об орхидеях был бы неполным, если бы мы не остановились
хотя бы коротко на особенностях процесса опыления. В описываемом семействе
оно необычайно своеобразно, что всегда привлекало внимание исследователей.
Первым, кто этот процесс подробно изучил с эволюционных позиций, был Чарлз
Дарвин. Высокая специализация процесса перекрестного опыления, характерная
для орхидей, служит весомым доказательством того, что они представляют
собой вершину эволюции однодольных растений.
Многолетний опыт наблюдений за природными орхидными убедительно свидетельствует, что они очень хорошо приспособились к тесному общению с определенными видами или близкими группами насекомых. Не исключена и более узкая специализация, когда та или другая орхидея плодотворно посещаются только одним видом опылителя. Очевидно, это не лучший вид гарантии продолжения рода. Чаще один вид насекомого посещает несколько видов растений или несколько видов насекомых — один вид растения. Это связано с тем, что насекомые вынуждены искать и другие виды растений, так как период цветения одного вида растения обычно короче, чем период, когда насекомое нуждается в пище. Чем более тесно связаны цветок и опылитель, тем полезнее для каждого из них эта связь. Однако здесь и потенциальная слабость этого союза — если один из партнеров исчезнет, то погибнет и второй, либо второй должен выработать какие-то новые приспособления, компенсирующие отсутствие партнера. Например, офрис пчелоносная отсутствие партнера компенсировала выработкой способности к самоопылению [Дарвин, 1950]. Насекомые посещают цветки орхидей, движимые самыми разными желаниями. Естественно, ради пищи, которой служат нектар и пыльца. В околоцветнике ими откладываются яички, имеются и сексуальные причины. Окраска, оригинальная форма, аромат орхидей — все это наряду с различными вспомогательными устройствами привлекает насекомых. В тропических странах опылителями становятся не только представители насекомых, но и птицы, тоже не менее успешно способствующие перекрестному опылению. Выделяющиеся в темноте снежно-белые венчики орхидей хорошо заметны для ночных бабочек. Птицы предпочитают красные тона, а мясные мухи — грязно-фиолетовые или коричнево-красные оттенки [Гладкова, 1982]). Запахи играют не менее важную роль в привлечении опылителей. Тем более что они бесконечны по своей гамме: от зловония, напоминающего о гнилом мясе, до букета нежнейших оттенков, ласкающих обоняние как самые изысканные духи. Нередко последним достоинством обладают внешне малопривлекательные зеленовато-белые цветки. Установлено в настоящее время, что половину переносчиков пыльцы составляют перепончатокрылые насекомые — пчелы, осы, шмели. Второе место по численности занимают чешуекрылые (различные бабочки). Несколько меньше—двукрылые (мухи, оводы, комары). Вклад птиц в опыление невелик, в основном это колибри, сами напоминающие красивые цветы. Наиболее замечательными приспособлениями орхидей к опылению можно считать
использование ими непищевых инстинктов насекомых (сексуальных, защитных)
и широкое привлечение к опылению самцов пчел, ос. Объясняют такую своеобразную
адаптацию растений тем, что орхидные появились сравнительно поздно, когда
уже существовало множество растений, снабжавших опылителей пищей (пыльцой
и нектаром), и орхидеи вынуждены были искать новый, непроторенный путь
обеспечения опыления.
Среди множества особенностей механизма опыления у орхидей обращает на себя внимание одно любопытное свойство. Оно для них универсально и заключается в длительном цветении, что касается как отдельных цветков, так и целых соцветий. Венерин башмачок обыкновенный в ожидании опыления может не вянуть до 3 нед, но сразу завядает, если к рыльцу прилипает пыльца. Ятрышники держатся 7 — 10 дней, а некоторые тропические орхидеи — 2—3 мес. В большинстве случаев соцветие распускается не все сразу, а постепенно снизу вверх и имеет свежий вид в течение нескольких недель, а то и месяцев. Очень важно, что у орхидных даже единственное посещение насекомого-опылителя становится достаточным, чтобы произошло завязывание очень многих семян. Пыльцевые зерна орхидей, во всяком случае у большинства из них, связаны друг с другом и не могут разноситься ветром. Поэтому опыление может происходить лишь с помощью животных — насекомыми или птицами. Как пищевой фактор пыльца большого значения не имеет. Она компактно упакована в поллинии, попадающие па рыльце цветка целиком. При этом пыльцевые зерна не пропадают впустую, а сразу оплодотворяют огромное количество яйцеклеток, из которых развиваются тысячи семян. Яркий и броский околоцветник служит рекламой другого ценного продукта — нектара, предлагаемого насекомым растением взамен за их услуги. Иногда сладкий сон располагается открыто, выделяясь в бороздках губы, как, например, у тайника овального. Чаще нектар накапливается в шпорце, отходящем от основания губы или, что реже, от другого листочка околоцветника. Разная длина шпорца предполагает и соответствующие размеры хоботков опылителей. У тропической орхидеи из рода ангрекум шпорец достигает в длину 30 см, и можно представить себе насколько велико должно быть насекомое. Известно, что такой длины достигает свернутый в спираль хоботок некоторых тропических бабочек. Однако далеко не всегда насекомые находят в цветках желаемую пищу. Орхидеи способны ловко их обманывать. В шпорцах нередко отсутствует нектар, а желтые пятна на губе, так похожие на пыльники, наполненные пыльцой, оказываются скоплениями окрашенных волосков. Привлекательные чаши, наполненные вкусной жидкостью, оборачиваются хитрыми ловушками. Одним словом, цветки стремятся показать, что в них находится немало пищи или же присутствует половой партнер, и насекомое доверчиво летит на хитрую приманку. Конечно, желания насекомых не удовлетворяются, но перекрестное опыление они совершают. Хорошо, что природа знает меру и видов-имитаторов не так уж много. Во всяком случае, они не превышают по численности те растения, которые служат им моделью и обладают не призрачными, а настоящими богатствами, В противном случае насекомые, убедившись в постоянном обмане, перестали бы посещать цветки таких орхидей. Наблюдая столь оригинальное природное явление, некоторые исследователи пытаются его оценить с позиций человеческих чувств. Они говорят о «дипломатии» орхидей, о «сообразительности» и «остроумии» и даже о «безнравственности» орхидей. Это служит своеобразным дополнением к обширной литературе, посвященной особенностям их опыления. Кстати, все сказанное целесообразно, видимо, проиллюстрировать примерами. Одной из самых крупных и красивых наземных орхидей заслуженно считается
венерин башмачок обыкновенный. Цветок его представляет настоящую ловушку,
и насекомое, попадающее внутрь его, вынуждено произвести опыление. Крупная
желтая губа имеет форму туфельки с завернутым внутрь краем. Вдобавок сверху
она закрыта щитковидным стаминодием (бесплодной тычинкой),
и рыльце расположено под ним. Его поверхность, воспринимающая пыльцу, направлена
вниз, к «туфельке», а по обеим сторонам от рыльца расположено по плодущей
тычинке. Одиночные пчелы, особенно самки из рода андрена, вползают в губу
цветка и после ее обследования начинают искать выход. Ограниченное пространство
не позволяет им взлететь, а завернутые края губы, гладкая внутренняя поверхность
губы и заслон из стаминодия не позволяют выползти прямым путем на волю.
В задней стенке губы имеются участки ткани, образованные клетками без пигментации,
хорошо пропускающими свет (так называемые «окна»). Насекомое принимает
их за выход и ползет в указанном ему направлении, по не добивается успеха.
Повернувшись, оно уже видит настоящий выход наружу — два узких отверстия
справа и слева от колонки. Выбираясь из цветка на свободу, насекомое сначала
встречает на своем пути выпуклое рыльце. Грудь насекомого проходит непосредственно
над рыльцем, и о него насекомое очищает принесенную пыльцу. Насекомое поднимается
все выше и выше ближе к свету и обнаруживает новое препятствие — выход
сужен, так как пыльник частично загораживает отверстие. Насекомое с трудом
протискивается мимо пыльника, и большая порция пыльцы (у башмачков она
не склеена в поллинии) приклеивается к груди насекомого.
Для мелких или ослабленных насекомых последний рубеж может оказаться роковым. Иногда они приклеиваются к пыльнику и не в состоянии от него оторваться, оставаясь здесь навсегда, закупоривая вход в цветок. Что же касается нашей пчелы, которой удалось выбраться, то она, уставшая, голодная, нередко еще и мокрая (так как выкупалась в дождевой воде, скопившейся на дне «туфельки»), некоторое время даже не в состоянии нормально двигаться. Но отдохнув, пчела летит на другие цветки. Таким образом, прекрасные башмачки тоже относятся к категории «обманщиков», привлекая насекомых нежным запахом и ярким цветом губы и стаминодия, якобы свидетельствующими о нектаре (рис. 8) [Фегри, Пейл, 1982] Для многих наших орхидей с хорошо выраженной плоской губой, играющей
роль «посадочной площадки», характерна иная схема опыления, не менее оригинальная,
которую мы рассмотрим на примере ятрышника мужского. Завязь у этого вида
перекручена таким образом, что губа занимает нижнее положение и может выполнять
роль посадочной площадки. Губа имеет короткий шпорец. Когда насекомое,
севшее на губу, просовывает голову в полость, ведущую в шпорец, оно неизбежно
наталкивается на выступающий в этот проход «клювик» (неплодущая часть рыльца,
превратившаяся в мешочек, в котором находятся липкие подушечки поллинариев).
От энергичного прикосновения конец клювика сразу расщепляется, а эластичный
карманчик отгибается, обнажая 2 прилипальца, которые касаются головы насекомого
и, быстро твердея, крепко приклеиваются к нему в виде двух рожек. Когда
насекомое извлекает голову из цветка, оно вытаскивает поллиний (или оба),
а эластичный мешочек при этом захлопывается снова. Прикрепившиеся поллинии
сначала стоят вертикально, а примерно через 30, о поллинарий благодаря
асимметричному высыханию изгибается под углом 90°, и при посещении насекомым
следующего цветка поллинии попадают точно на его рыльце. Поверхность рыльца
липкая, и поллиний остается на ней. Пчела улетает с остатками хвостиков
поллиниев на голове как доказательством того, что работа успешно выполнена.
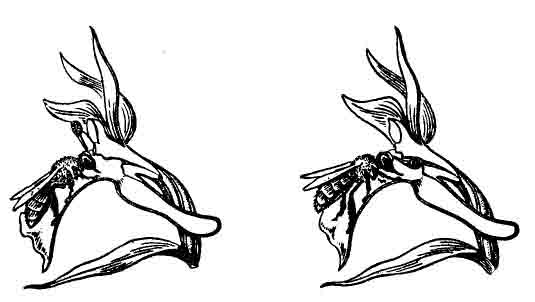
К этой же категории относится и пыльцеголовник красный. Соцветие его состоит из 4—10 бросающихся в глаза розовых довольно крупных цветков, полностью лишенных нектара или какой-либо другой пищи, необходимой для насекомых. В связи с этим он вынужден завлекать насекомых, подражая видам колокольчиков, которые охотно посещаются насекомыми. Установлено, что опыляют пыльцеголовник самцы двух видов одиночных пчел. Самки этих видов собирают пыльцу исключительно с колокольчиков. Проведенный колориметрический анализ окраски цветков пыльцеголовника доказал сходство их «в глазах» пчел (как известно, насекомые воспринимают цвет иначе, чем люди). Химический состав цветков пыльцеголовника и колокольчика персиколистного (одного из самых крупноцветковых и красивых наших колокольчиков) также оказался сходным [Dafni, Ivri, 1981]. Еще более замечательный способ опыления наблюдается у орхидей рода офрис.
Цветки этих необычных и редких растений часто напоминают насекомых — мух,
пчел, шмелей, оводов. От этого сходства и происходят их видовые названия
— офрис насекомоносная, мухоносная, шмелецветковая и т. д. У цветков офрис
насекомоносной, например, губа похожа на тело насекомого со сложенными
крыльями, блеск которых имитируют серо-голубые полоски на губе. Боковые
листочки околоцветника имеют вид усиков, а расположенные в основании губы
ложные нектарники напоминают глаза насекомого. В целом цветок удивительно
удачно имитирует облик самки того или иного вида насекомых. Предполагают
также и сходство запаха цветка и насекомого. Этому, наверно, трудно поверить,
однако изучение химического состава выделений цветка и насекомого подтвердило,
что они сходны по своему составу. Насекомые находят такой цветок, даже
если он спрятан в темном помещении. Собственно, особых чудес здесь нет.
Самцы ряда перепончатокрылых (андрена, эвцера, антофора и пр.) появляются
после перезимовки несколько раньше самок. Побуждаемые мощным сексуальным
инстинктом, они летят к цветкам офрис, похожим по форме и запаху на самок.
В результате импульсивных движений тела поллинии прилипают к голове и;
брюшку насекомого. Для удовлетворения полового инстинкта посещения одного
цветка оказывается недостаточно, и насекомое посещает еще несколько цветков,
осуществляя тем самым перекрестное опыление. Такая приспособленность офрис
к специфическим опылителям могла возникнуть, видимо, в результате длительной
сопряженной эволюции орхидных и насекомых (рис. 9).
Наконец, необходимо упомянуть еще одну группу орхидей, которой свойственно
самоопыление. Число входящих в нее видов может колебаться. Если по какой-либо
причине исчезает узкоспециализированный опылитель, то растение вынуждено
заботиться само о своем потомстве, о продолжении своего рода, переходя
к самоопылению. Предполагают, что самоопыление офрис пчелоносной вызвано
именно этой причиной. Известны и закрытые (клейстогамные) цветки, которые
развиваются нередко подземно, самоопыляются, не раскрываясь, и дают семена.
Иногда такое явление отмечают даже у нашей гнездовки
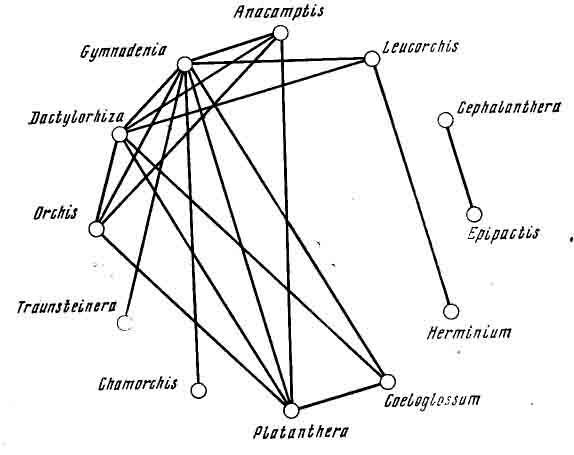
Семейство орхидных появилось одним из последних в растительном мире нашей планеты. Об относительной молодости этого семейства и о продолжении процесса видообразования свидетельствует та легкость, с которой они образуют гибриды. Впервые были обнаружены гибриды именно у европейских орхидей. В Альпах был найден гибрид между нигрителлой черной и кокушником комарниковым. В настоящее время известно значительное число гибридов орхидей как между видами, так и между родами. Иногда даже встречаются более сложные гибриды, образованные тремя видами. Чешские ученые Ф. Прохазка и В. Величек [Ргосhazka, Velisek, 1983] приводят
схемы возможных гпбридов между отдельными видами и родами (рис. 10), из
которых следует, что род пальчатокоренник может гибридизировать с 6 родами
(ятрышником, кокушником, анакамптисом, псевдорхисом, любкой, пололепестником),
род кокушник — с 8 родами, роды ятрышник и анакамптис — с 4 родами, любка
— с 5 родами. Однако роды пыльцеголовник и дремлик, близкие по многим признакам,
гибридизируют только между собой, не вступая в связи с другими родами.
Особенно энергично гибридизируют между собой виды рода пальчатокоренник
(поэтому, видимо, так запутана систематика этого рода). Своеобразными «чемпионами»
среди видов этого рода являются пальчатокоренники Фукса и майский, которые
вступают в гибридизацию с 6 другими видами своего рода, лишь немного им
уступают пальчатокоренники пятнистый и мясо-красный, гибридизирующие с
5 видами своего рода.
Такое удивительное свойство орхидей таит в себе необычайно широкие возможности. Оно давно используется при разведении тропических видов в оранжереях. Создаются гибриды поистине сказочного вида. Гибриды наших дикорастущих орхидей также нередко исключительно красивы по цвету и форме. Особенно часто находят гибридные формы орхидей в горных районах — в Крыму, на Кавказе, в Карпатах. Нельзя исключить, что со временем гибридные формы наших дикорастущих орхидей займут свое место в садах и парках. Орхидеи нашей страны/М. Г. Вахрамеева, Л. В. Денисова, С. В. Никитина, С. К. Самсонов.— М.: Наука, 1991.— 224 с |
| Посещений страницы: 14618 |
|
|
|
|
|
|